
Jądro komórkowe

2. jąderko
3. karioplazma
4. chromatyna 4a. Heterochromatyna 4b. Euchromatyna
5. Rybosomy
6. Pory jądrowe
Jądra komórkowego lub jądro ( Łacińskiej jądro „rdzeń”) jest najczęściej okrągły organelli z w eukariotycznej komórce znajduje się w cytoplazmie , które zawiera genetyczny materiał. W jądrze jądrowym lub karyo (w starożytnej grece κάρυον káryon „jądro”) wyrażane jest odniesienie do jądra komórkowego, które nazywa się genomem jądrowym (w przeciwieństwie do tego w organellach obwodowych), na przykład także karyoma . Nauka o jądrze nazywana jest również kariologią .
opis
Jądro komórkowe jest główną cechą odróżniającą eukariota (istoty żywe z ograniczonym jądrem komórkowym) i prokarionty (istoty żywe bez ograniczonego jądra komórkowego, tj. bakterie i archeony ). Zawiera większość materiału genetycznego komórek eukariotycznych w postaci kilku chromosomów (jądrowego DNA lub jądrowego DNA). Inne geny można znaleźć w mitochondriach i wyjątkowo w hydrogenosomach , a także w glonach i roślinach lądowych w chloroplastach i innych plastydach . Większość komórek zawiera dokładnie jedno jądro. Są jednak wyjątki ( syncytium ); na przykład miotuby, które powstają w wyniku fuzji mioblastów , zawierają wiele jąder. Samo jądro jest oddzielone od otaczającej cytoplazmy otoczką jądrową lub błoną jądrową, która służy do ochrony materiału genetycznego i regulacji transportu substancji między nukleoplazmą a cytoplazmą . W zarodkach muszek owocowych jądra dzielą się bardzo szybko, nie tworząc początkowo oddzielających się błon komórkowych . Dojrzałe erytrocyty wśród ssaków nie zawierają rdzeń anymore, jest odpychany podczas dojrzewania.
Ważnymi procesami zachodzącymi w jądrze komórkowym są replikacja DNA (duplikacja materiału genetycznego w postaci DNA ) oraz transkrypcja (utworzenie kopii mRNA danego odcinka DNA, która często, ale nie zawsze, odpowiada genowi ).
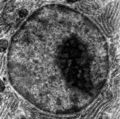
Obraz jądra komórkowego z mikroskopu elektronowego

Jądro komórkowe z Medicago truncatula

Cebula ( Allium cepa ) jądro, 400 ×, barwione jodem

.png)

budowa

1. Jąderko (ciało jądrowe)
2. Jądro komórkowe (jądro komórkowe)
3. Rybosomy
4. Pęcherzyk
5. Szorstki (ziarnisty) ER (ergastoplazma)
6. Aparat Golgiego
7. Cytoszkielet
8. Gładki (agranularny) ) ER
9. mitochondria
10. lizosom
11. cytoplazma (z cytozolem i cytoszkieletem )
12. peroksysomy
13. centriole
14 błona komórkowa
Jądro komórkowe, które u ssaków zwykle ma średnicę od 5 do 16 µm , jest organellą komórki, którą najłatwiej zobaczyć pod mikroskopem. Jest on ograniczony otoczką jądrową , składającą się z dwóch błon biologicznych, wewnętrznej i zewnętrznej błony jądrowej, które otaczają tzw. cysternę okołojądrową (szerokość 10–15 nm, wzmocniona mikrowłókienkami – grubość od 2 do 3 nm). Całkowita grubość otoczki jądrowej wynosi około 35 nm Zewnętrzna błona jądrowa łączy się płynnie z szorstką siateczką endoplazmatyczną (rER) i, w ten sposób, ma również rybosomy na swojej powierzchni. Wewnętrzna błona jądrowa graniczy z „filcem” o szerokości 20–100 nm, blaszką jądrową (Lamina fibrosa nuclei), która składa się z lamin , grupy włókien pośrednich , które podtrzymują jądro komórkowe i oddzielają błonę wewnętrzną od chromatyny jądro komórkowe.
Jądra komórkowe mogą wyglądać bardzo różnie w zależności od typu komórki. Najczęściej są kuliste lub owalne. W niektórych komórkach wyglądają bardziej jak poroże. Czasami jądro można podzielić na sekcje przypominające węzły, jak to ma miejsce w przypadku jądra w kształcie różańca trąbek . W granulocyty wśród ssaków również zawierać klapowane jąder.
Ze względu na zawarte w otoczce jądrowej pory jądrowe , które pokrywają ok. 25% powierzchni, następuje aktywna wymiana substancji (np. rRNA lub mRNA) między jądrem a osoczem komórkowym , kontrolowana przez kompleks porów jądrowych . Białka regulatorowe przechodzą z cytoplazmy do jądra komórkowego, produkty transkrypcji, takie jak mRNA, są eksportowane z jądra do osocza w celu syntezy białek , która odbywa się na rybosomach cytoplazmy. Płyn w jądrze znany jest również jako karioplazma . Jądra komórkowe można uwydatnić pod mikroskopem świetlnym poprzez barwienie DNA , np . B. metodą barwienia Feulgena , metodą Giemsy lub barwnikami fluorescencyjnymi, takimi jak DAPI .
Obecność macierzy rdzenia została po raz pierwszy zasugerowana w latach 70. XX wieku. Jednak ich istnienie wciąż budzi kontrowersje.
Materiał genetyczny komórki w jądrze komórkowym znajduje się w chromosomach , tj. H. kilka nici DNA upakowanych w chromatynie, które zawierają nie tylko DNA, ale także białka, takie jak histony . Oprócz histonów istnieją inne białka rdzeniowe, takie jak B. Polimerazy DNA i polimerazy RNA , inne czynniki transkrypcyjne i kwasy rybonukleinowe w jądrze.

Ciała jądrowe
Za pomocą mikroskopu świetlnego można zobaczyć jedną lub więcej zaokrąglonych struktur w wielu jądrach komórkowych, ciałkach lub jąderkach. Zawierają geny rybosomalnego RNA (rRNA). W tym miejscu powstają podjednostki rybosomów , które wchodzą do cytoplazmy przez pory jądrowe. Jąderka zawierają tylko niewielkie stężenia DNA w porównaniu z resztą jądra, a zamiast tego zawierają więcej RNA. Inne „ciałka” jądra komórkowego można uwidocznić tylko przy użyciu specjalnych technik barwienia, takich jak barwienie przeciwciał . Funkcja tych ciał jest w większości nadal nieznana. Należą do nich np. „ plamki ” (zbiór czynników wymaganych do splicingu ), korpusy Cajala czy korpusy PML . Przestrzenna separacja różnych składników niektórych ciał jądrowych mogłaby umożliwić bardziej efektywne przeprowadzanie interakcji molekularnych w ekstremalnie zatłoczonym środowisku w jądrze. .
| Przyjęta funkcja | Komponenty centralne | Typowy rozmiar (μm) | Typowa liczba | |
|---|---|---|---|---|
| Jądro | Produkcja rybosomów poprzez transkrypcję i przetwarzanie rRNA oraz budowanie podjednostek rybosomalnych. Odgrywa rolę w modyfikacji i produkcji innych jądrowych RNA i RNP. Reguluje przebieg cyklu komórkowego poprzez sekwestrację i modyfikację wielu białek. | RNA pol I. | 3-8 0,5-8 |
1-4 |
| Plamki jądrowe (ICG) | Przechowywanie, montaż i modyfikacja czynników splicingowych. | Czynniki splicingowe pre-mRNA, SRSF2, SRSF1, jabłczan1 | 2-3 0,5-1,8 |
20-50 |
| Ciała stresu jądrowego | Regulacja transkrypcji i splicingu pod wpływem stresu.
Zawiera niekodujące RNA Satellite III. Precyzyjna funkcja nie została jeszcze zdefiniowana. |
HSF1, HAP | 1-2 0,3-3,0 |
2-6 2-10 |
| Ciało locus histonów (HLB) | Synteza genów histonowych | NPAT, FLASH, U7 snRNP | 0,2-1,2 | 2-4 |
| Ciała Cajala / Klejnoty | Produkcja, dojrzewanie i recykling SNRNP. Odgrywa również rolę w montażu telomerazy i regulacji wydłużenia telomerazy. | Zwój, SMN | 0,2-1,5 0,1-2,0 |
0-10 |
| Ciało PML | Regulacja stabilności genomu, naprawa DNA, kontrola transkrypcji, obrona wirusów | Białko PML | 0,1-1,0 0,3-1,0 |
10-30 |
| Paraspeckle | Regulacja mRNA, przetwarzanie RNA | NEAT1 / MENε / βncRNA PSP1, p54nrb / NONO | 0,2-1 0,5 |
2-20 10-20 |
| Przedział okołojądrowy | Regulacja potranskrypcyjna podzbioru RNA pol III w komórkach nowotworowych. | PTB, CUGBP | 0,2-1,0 | 1-2 1-4 |
| Korpus z Polycomb (PcG) | Kompleksy Polycomb Group (PcG) są zaangażowane w represję genów poprzez epigenetyczną modyfikację chromatyny i regulację podstawowej organizacji ich docelowych genów. | Bmi1, Pc2 | 0,3-1,0 | 12-16 |
| Domena OPT | Wzbogacony czynnikami transkrypcyjnymi Oct1 i PTF. Częściowa kolokalizacja z miejscami transkrypcji. Wykrywany głównie w późnej fazie G1. Rozpada się, gdy transkrypcja jest zahamowana. | 1,0-1,5 | 1-3 | |
| Cząstki jądrowe Sam68 | Koncentruje białka Sam68 i Sam68-podobne SLM-1 i SLM-2. Rozpada się, gdy transkrypcja jest zahamowana. Najprawdopodobniej wzbogacony RNA. | 0,6-1,0 | 1-5 | |
| ciało dekolt | Wzbogacony czynnikami rozszczepiania CstF 64 kDa i CPSF 100 kDa oraz białkiem DEAD box DDX1. Jest rozpoznawana głównie w fazie S i nie ma na nią wpływu inhibicja transkrypcji. | 0,2-1,0 | 1-4 | |
| Klastosomy | Ciała te koncentrują substraty białkowe w celu rozpadu proteasomów. Rozpoznawany głównie, gdy stymulowana jest aktywność proteasomu. Rozkłada się po inhibicji proteasomów. Tworzy się w odpowiedzi na bodźce aktywujące proteolizę zależną od proteasomu. | Proteasomy 19S, 20S | 0,2-1,2 | 0-3 |
| SUMO ciało | Wzbogacony o SUMO-1 i enzym sprzęgający SUMO Ubc9. Koncentruje czynniki transkrypcyjne pCREB, CBP, c-Jun. | 1-3 | 1-3 | |
| PIKA | nieznany (polimorficzna interfaza asocjacji kariosomalnej) |
Układ chromosomów

Podczas interfazy chromosomy zajmują określone obszary w jądrze komórkowym, terytoria chromosomów . Ich istnienie po raz pierwszy zasugerowali Carl Rabl (1885) i Theodor Boveri (1909), a bezpośrednie wykrycie było możliwe dopiero w 1985 roku za pomocą hybrydyzacji fluorescencyjnej in situ .
Rozkład chromatyny, a tym samym i chromosomów w jądrze komórkowym, wydaje się na pierwszy rzut oka przypadkowy: ułożenie chromosomów zmienia się z jądra na jądro, sąsiedzi w jednym mogą być daleko od siebie w następnym. Jednak od lat 90. znaleziono pewne zasady porządku. Replikacja DNA nie zachodzi jednolicie podczas fazy S , ale raczej wcześniej w niektórych częściach chromosomów, a później w innych. Wczesna lub późna replikacja to właściwości, które są stałe dla wszystkich odcinków chromosomów w danym typie komórki. Okazało się, że wczesne obszary replikacji znajdują się głównie wewnątrz jądra, podczas gdy obszary późnej replikacji znajdują się głównie na otoczce jądrowej i wokół jąder. Odnośnie rozmieszczenia terytoriów chromosomowych w jądrze komórkowym zaobserwowano, że chromosomy o dużej gęstości genów są korzystnie zlokalizowane w środku jądra, podczas gdy chromosomy o małej gęstości genów częściej znajdują się na obrzeżach. W przypadku niektórych typów komórek opisano również, że małe chromosomy znajdują się bardziej pośrodku, podczas gdy duże znajdują się na zewnątrz. Oba motywy są ze sobą kompatybilne.
Podział podstawowy
W mitozie i mejozie , rodzajach podziału jądra, które występują w komórkach eukariotycznych, jądro komórkowe tymczasowo zanika, ponieważ otoczka jądrowa jest rozpuszczana podczas procesu podziału. Chociaż chromosomy nie tworzą żadnych mikroskopijnych granic świetlnych w interfazie , kondensują się w zwarte chromosomy metafazowe w celu podziału jądra. W tej formie transportu materiał genetyczny jest rozprowadzany do komórek potomnych. Po podziale otoczki jądrowe ponownie tworzą się wokół chromosomów komórek potomnych i chromosomy ponownie ulegają dekondensacji.
Historia badań


Poniżej: Przedstawienie fałszywych kolorów wszystkich terytoriów chromosomowych widocznych w tej płaszczyźnie ogniskowej, zgodnie z klasyfikacją komputerową.
Jądro komórkowe jest pierwszym organellą odkrytą w komórce. Najstarszy zachowany rysunek pochodzi od wczesnego mikroskopijnego Antoniego van Leeuwenhoeka (1632–1723). Ten zbadano czerwone krwinki z łososia i opisane A „światło”, do jądra komórki. W przeciwieństwie do czerwonych krwinek ssaków, te innych kręgowców mają jądra komórkowe. Kolejną wzmiankę poczynił w 1804 r. Franz Andreas Bauer . W 1831 r. szkocki botanik Robert Brown opisał jądro jako „areola” w wykładzie wygłoszonym dla Towarzystwa Linneusza w Londynie . Nie wspomniał o możliwych znaczeniach. Po raz pierwszy zasugerował to Matthias Schleiden w 1838 roku , a mianowicie, że odgrywa ona rolę w tworzeniu komórki. Dlatego Schleiden wprowadził nazwę „Cytoblast” (konstruktor komórek). Powiedział, że zaobserwował, że na tych cytoblastach tworzą się nowe komórki. Franz Julius Ferdinand Meyen zdecydowanie zaprzeczył poglądowi, że „jądro komórkowe tworzy samą komórkę”. Wcześniej opisał, że komórki mnożą się przez dzielenie. Jednak Meyen był również zdania, że wiele komórek nie ma jądra komórkowego. Ideę tworzenia nowych komórek przezwyciężyły dopiero prace Roberta Remaka (1852) i Rudolfa Virchowa ( Omnis cellula e cellula , 1855), którzy agresywnie opowiadali się za nową teorią wyłącznego tworzenia komórek z komórek. Funkcja jądra komórkowego pozostała niewyjaśniona.
Christian Gottfried Ehrenberg po raz pierwszy zaobserwował podział jądra komórkowego w 1838 roku i zwrócił uwagę na jego rolę.
1876–1878 Oscar Hertwig opublikował kilka badań dotyczących procesów zapłodnienia jaja jeżowca morskiego , z których wyszło, że jądro komórkowe plemnika przenika do komórki jajowej i tam łączy się z jądrem komórkowym komórki jajowej. Po raz pierwszy twierdzono, że dana osoba rozwija się z (pojedynczej) komórki jądrzastej. To sprzeczne Ernst Haeckel pogląd, jakoby cała plemienna historia będzie się powtarzać w czasie rozwoju embrionalnego, w szczególności pojawienie się pierwszej komórki jądrzastych z «Monerula», o masie structureless pierwotnej śluzu. Potrzeba jądra plemnika do zapłodnienia była więc przez długi czas kontrowersyjna. Hertwig potwierdził jednak swoje odkrycia w innych grupach zwierząt, m.in. B. Płazy i mięczaki . Eduard Strasburger doszedł do tego samego wniosku w sprawie roślin (1884). To utorowało drogę rdzeniowi do odegrania ważnej roli w dziedziczeniu. Już w 1873 roku August Weismann postulował równoważność matczynych i ojcowskich komórek rozrodczych do dziedziczenia. Rola jądra w tym miejscu ujawniła się dopiero później, po opisaniu mitozy i ponownym odkryciu zasad Mendla na początku XX wieku : rozwinięto chromosomową teorię dziedziczenia (patrz tam i chromosom ).
1874 wyizolował Friedricha Mieschera z jąder komórkowych, substancję, którą nazwał nukleinową (patrz historia odkrycia w artykule DNA ). Dopiero kilka lat później dodano inne składniki, takie jak histony i adenina ( Albrecht Kossel ). Dopiero od XXI wieku możliwe jest określenie genomu , transkryptomu lub, za pomocą spektrometrii mas , proteomu komórki w określonym momencie w celu uzyskania pełnego obrazu tego układu.
Zobacz też
literatura
- T. Cremer: Od teorii komórek do teorii chromosomów . Springer Verlag, Berlin i in., 1985, ISBN 3-540-13987-7 (online) (o historii; PDF; 6,1 MB)
linki internetowe
Indywidualne dowody
- ↑ Shinichi Nakagawa, Tomohiro Yamazaki, Tetsuro Hirose: Molekularna dysekcja jądrowych paraspek: ku zrozumieniu rodzącego się świata środowiska RNP . W: Otwarta Biologia . taśma 8 , nie. 10 października 2018 r., ISSN 2046-2441 , s. 180150 , doi : 10.1098/ rsob.180150 , PMID 30355755 , PMC 6223218 (darmowy pełny tekst).
- ↑ a b c d e f g h i j k l m Miroslav Dundr, Tom Misteli: Biogeneza ciał jądrowych . W: Cold Spring Harbor Perspectives in Biology . taśma 2 , nie. 12 grudnia 2010, ISSN 1943-0264 , doi : 10.1101 / cshperspect.a000711 , PMID 21068152 , PMC 2982170 (pełny tekst dowolny).
- ^ B c d Maria Carmo-Fonseca Maria T. Berciano Miguel Lafarga: Zbiorniki sierocego jądrowej . W: Cold Spring Harbor Perspectives in Biology . taśma 2 , nie. 9 września 2010, ISSN 1943-0264 , doi : 10.1101 / cshperspect.a000703 , PMID 20610547 , PMC 2926751 (wolny pełny tekst).
- ↑ a b c d e f g h i j k l m n o Yuntao S. Mao, Bin Zhang, David L. Spector: Biogeneza i funkcja ciał jądrowych . W: Trendy w genetyce . taśma 27 , nie. 8 , sierpień 2011, s. 295-306 , doi : 10.1016 / j.tig.2011.05.006 , PMID 21680045 , PMC 3144265 (pełny tekst dowolny) - ( elsvier.com [dostęp 2 maja 2019 r.]).
- ↑ Carmelo Ferrai, Inês Jesus de Castro, Liron Lavitas, Mita Chotalia, Ana Pombo: Pozycjonowanie genów . W: Cold Spring Harbor Perspectives in Biology . taśma 2 , nie. 6 czerwca 2010, ISSN 1943-0264 , doi : 10.1101 / cshperspect.a000588 , PMID 20484389 , PMC 2869523 (pełny tekst dowolny).
- ^ Kompartmentalizacja w jądrze: Odkrycie nowego regionu podjądrowego . W: The Journal of Cell Biology . taśma 115 , nie. 4 , 2 listopada 1991, ISSN 0021-9525 , s. 919-931 , PMID 1955462 , PMC 2289954 (wolny pełny tekst).
- ↑ Schardin i in. : Specyficzne barwienie ludzkich chromosomów w hybrydowych liniach komórkowych chomika chińskiego i człowieka wskazuje na obszary międzyfazowego chromosomu. Szum. Genet. 71: 281, 1985. PMID 2416668 .
- ↑ Manuelidis: Poszczególne domeny chromosomów międzyfazowych ujawnione przez hybrydyzację in situ . Szum. Genet. 71:288, 1985. PMID 3908288 .
- ↑ O'Keefe i in. : Dynamiczna organizacja replikacji DNA w jądrach komórek ssaków: przestrzennie i czasowo określona replikacja sekwencji alfa-satelitarnych DNA specyficznych dla chromosomu. J. Cell Biol 116: 1095, 1992. Streszczenie i pełny tekst w Journal of Cell Biology (angielski).
- ^ Cremer i Cremer : Terytoria chromosomów, architektura jądrowa i regulacja genów w komórkach ssaków. Nat Rev Genet 2: 292, 2001. PMID 11283701 .
- ^ Antoni van Leeuwenhoek Opera Omnia, zob. Arcana Naturae ope ścisła Microscopiorum Detecta, eksperymentis variis comprobata, Epistolis ad varios illustres viros. J. Arnold i Delphis, A. Beman, Leiden 1719-1730. Cytat z Dieter Gerlach: Historia mikroskopii. Verlag Harry Deutsch, Frankfurt nad Menem 2009, ISBN 978-3-8171-1781-9 .
- ^ H. Harris: Narodziny komórki . Wydawnictwo Uniwersytetu Yale, New Haven 1999.
- ^ Robert Brown: O narządach i sposobie zapłodnienia Orchidex i Asclepiadea . W: Różne prace botaniczne . taśma I , 1866, s. 511-514 .
- ↑ Bärbel Häcker: Chromosomy. W: Werner E. Gerabek , Bernhard D. Haage, Gundolf Keil , Wolfgang Wegner (red.): Enzyklopädie Mediizingeschichte. De Gruyter, Berlin / Nowy Jork 2005, ISBN 3-11-015714-4 , s. 261 f.
- ↑ W. Waldeyer : O kariokinezie i jej związku z procesami zapłodnienia . W: Archiwum anatomii mikroskopowej . taśma 32 , nie. 1 , 1888, s. 1–122 , doi : 10.1007 / BF02956988 ( PDF ).